Коёкина О.И.
Научный центр исследований сознания, Москва, e-mail: Адрес электронной почты защищен от спам-ботов. Для просмотра адреса в вашем браузере должен быть включен Javascript.
Целью работы являлись поиск и определение способа обработки сенсорных сигналов в первичных сенсорных зонах коры мозга отдельных видов восприятия: аудиального, визуального, кинестетического. Показано, что альфа-ритм специфичен в ограниченных частотных диапазонах по отношению к модальности сенсорных сигналов. Альфа-ритм частотой 7–9 Гц относится к соматосенсорному, 9–12 Гц – к зрительному, 12–13 Гц и 7–9 Гц – к слуховому восприятию. Эквивалентные дипольные источники альфа-ритма в первичной сенсорной зоне коры, по ходу афферентных путей, в структурах лимбической области и восходящей неспецифической активации объединяются в систему каналов передачи сенсорных сигналов отдельной модальности в едином частотном диапазоне. Частотная специфичность каналов передачи сенсорных сигналов ля каждой модальности может быть обусловлена стробированием – выделением интервала на шкале частот для увеличения вероятности обнаружения полезных сигналов на фоне помех. Подтверждено предположение эфаптической передачи в средах мозга сигналов разных сенсорных модальностей в область доминирующего центра. Способы самоорганизации работы мозга в виде альфа-стробирования и альфа-сканирования активности в отдельных структурах могут иметь дальнейшее развитие при изучении свойств сознания и эмоций. Результаты исследований могут быть использованы в различных сферах деятельности человека, имеющих отношение к психологии, искусствоведению, педагогике, к творческим специальностям.
Ключевые слова: электроэнцефалограмма, сенсорный центр, альфа-ритм, частота альфа-ритма, альфасканирование, альфа-стробирование, эквивалентный дипольный источник
Интегральная деятельность мозга связана, прежде всего, с комплексными системами взаимодействия, определяемыми по разнообразным функциональным параметрам [1, 2]. В процессе нейрофизиологических исследований можно наблюдать как отдельные признаки взаимодействия в структурах мозга выстраиваются в алгоритмы выполнения тех задач, которые возникают
перед субъектом [3]. Целью работы являлись поиск и определение алгоритма альфастробирования сигналов в первичных сенсорных зонах коры мозга отдельных видов восприятия: зрительного, слухового, соматосенсорного. Конкретной целью начального этапа данного исследования являлось выявление особенностей волновых колебаний альфа-ритма, связанных с модальностью сенсорных сигналов.
Известно, что непосредственная передача внешних сигналов от органов восприятия к первичным сенсорным центрам коры мозга выполняется высокочастотными ритмами. Это показано методами вызванных потенциалов и смещением спектра ЭЭГ в высокочастотную область, включающую гамма- и бета-диапазоны ритмов [4, 5]. Однако значение в этом процессе колебаний альфа-диапазона ЭЭГ до сих пор остаётся дискуссионным. Представление о том, что альфа-ритм – это отображение сканирования зрительной коры, считывания с нее информации для последовательной передачи ее в другие корковые области было выдвинуто впервые в качестве идеи (Pitts, McCulloch, 1947). Позднее эта идея была подтверждена тщательными исследованиями альфа-ритма ЭЭГ как распространяющегося волнового процесса с одновременным смещением эквивалентного токового диполя альфа-волны [6, 7]. Важной особенностью упомянутых исследований было сопоставление модуляции альфа-волн световыми вспышками близкой частоты с возникновением зрительных образов, что подтверждало отношение альфа-ритма к сканированию зрительной информации в структурах первичного восприятия. При этом было показано также, что при совмещении локализации диполей альфа-волны с трехмерной
моделью головы, построенной по данным магнитно-резонансной томографии, наблюдается их передвижение преимущественно вдоль шпорной борозды (стриарная кора или поле 17). Следовательно, в полном объёме результаты сканирования альфа-волной были получены на примере активности первичного центра зрительного анализатора в коре мозга.
С учётом результатов вышеописанных исследований автором настоящей работы продолжено изучение процессов сканирования в первичных центрах сенсорного восприятия и одновременно поставлен вопрос, может ли иметь значение частотная специфичность альфа-волн при передаче сигналов разной модальности (визуальной, аудиальной, кинестетической).
Таким образом, в данной работе предполагалось, что критерием оценки поступления специфических сигналов в первичные зоны и показателем межзональных взаимодействий могут служить параметры спектральных характеристик альфа-ритма ЭЭГ.
На особенности частотного спектра альфа-ритма ЭЭГ первым обратил внимание Н. Винер [8]. Он особо отметил то, что преобладающая часть мощности спектра ритмов мозга сосредоточена в окрестности частоты 10 Гц. Определив в энцефалограмме конкретного человека крутое падение мощности на частоте 9,05 Гц, достаточно устойчивое во времени, он отметил, что резкая
линия смены частоты эквивалентна точным часам, обеспечивающим механизмы стробирования. Стробирование (англ. strobing, от strobe – посылать избирательные импульсы, от греч. strobos – кружение, беспорядочное движение), метод выделения некоторого интервала на временной оси, шкале частот и т.п. для увеличения вероятности обнаружения полезных сигналов на фоне помех [9].
При этом стробирование можно рассматривать как один из механизмов в развитии системы саморегуляции, так как позволяет осуществлять комбинирование действий в определяемых им интервалах времени.
Поэтому перед настоящим исследованием встала задача выяснить, может ли процесс стробирования, если он имеет место в альфа-диапазоне частот, обуславливать частотную специфичность для каждой отдельной зоны сенсорной модальности. Важно также понять и возможности взаимодействия между разномодальными зонами первичного восприятия с использованием эффекта стробирования. Поставленная задача может быть выполнена, если предусмотреть участие испытуемых с относительно равномерно развитыми типами восприятия. Для альфа-волн в каждой первичной зоне сенсорного восприятия наличие иной устойчивой доминирующей сенсорной модальности может влиять на поступление сигналов и их стробирование и сканирование. В исследовании необходимо было учитывать возможность возникновения увеличенного разброса данных при участии лиц с различными типами доминирующего восприятия.
Поэтому для получения более устойчивых результатов из группы испытуемых были исключены лица с преобладающим доминированием той или иной сенсорной модальности: визуалы, аудиалы, кинестетики. Таким образом, для выполнения поставленной задачи необходимо было ответить на следующие вопросы, имеющие непосредственное отношение к структуре исследования.
1. Какая частота альфа-ритма характерна для каждой зоны первичного восприятия?
2. Где располагаются источники происхождения альфа-ритма каждой корковой зоны первичного восприятия?
3. Как глубинные источники происхождения альфа-ритма проецируют активность в кору головного мозга?
Материалы и методы исследования
В исследовании участвовала группа добровольных испытуемых, занимающихся интеллектуальным трудом (30 чел., из них 17 женщин и 13 мужчин), в возрасте от 30 до 60 лет, не имеющих какой либо доминирующей перцептивной модальности, проверенной по методике С. Ефремцева [10]. Для получения ответов на выше поставленные вопросы 1–3 была проведена регистрация электроэнцефалограммы (ЭЭГ) в состоянии психической и мышечной релаксации с закрытыми глазами.
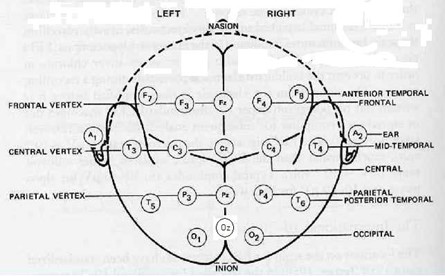
Рис. 1. Стандартное расположение электродов на поверхности головы по схеме 10/20
Регистрацию ЭЭГ проводили на 24-канальном нейровизоре NVX24 производства ООО «Медицинские компьютерные системы» с использованием стандартных монополярных отведений в соответствии с принятой международной схемой 10–20 [11].
Исследования проводились в стандартных условиях, одинаковых для всех испытуемых. Биопотенциалы мозга регистрировали при закрытых глазах в состоянии бодрствования с психической и мышечной релаксацией. Во внимание принимались только статистически достоверные результаты группового обследования.
Система обработки данных
Основная цель обработки данных регистрации ЭЭГ заключалась в топической оценке спектральных характеристик альфа-ритма и нейрокартирования с частотным шагом 1 Гц в пределах всего диапазона альфа-колебаний 7–13 Гц. Рассматривалась также локализация центров электрической активности, ответственных за происхождение отдельных частот альфа-ритма. Для этого использовали пакеты компьютерных программ, обеспечивающих спектральный анализ регистрируемых сигналов и топографическое картирование спектральных характеристик ЭЭГ [12]. В результате получали распределение показателей мощностичастотного спектра по отдельным областям на поверхности полушарий мозга. Использован статистический анализ данных для определения достоверности полученных как индивидуальных, так и групповых усреднённых результатов.
С целью выявления наиболее выраженных реакций вычисляли локализацию и распределение эквивалентных дипольных источников (ЭДИ) происхождения электрической активности в глубинных структурах мозга. Для этого определяли и отслеживали в объёме мозга человека локализацию ЭДИ, вычисляемых в каждый момент времени, равный дискретному интервалу отсчёта данных при вводе в компьютер. Программа использовалась для того, чтобы проследить возникновение центров электрической активности в структурах мозга [13].
Вопрос о соответствии локализации источников в глубинных структурах мозга той биоэлектрической активности, которая регистрировалась на поверхности головы, решался специальным алгоритмом, в котором рассматривались электрические свойства мозга как объёмного проводника [14, 15].
Результаты исследования и их обсуждение
Большинство людей имеют неустойчивые и низкие показатели доминанты, зависимые от ситуации в условиях меняющейся среды и психологического настроя. Испытуемые с устойчивыми и высокими показателями тестирования доминирующей перцептивной модальности (аудиалы, визуалы, кинестетики) по методике С. Ефремцева были исключены из группы исследования.
Поэтому у испытуемых в состоянии психической и мышечной релаксации наблюдалось равновесие активности первичных зон сенсорного восприятия. Оно выражалось в соответствии между частотой альфа-ритма и его максимальной мощностью в соответствующей зоне поступления первичных сенсорных сигналов.
В настоящее время анатомически и фукционально достаточно хорошо изучены области коры, которые отвечают за восприятие определённых ощущений, чувств, т.е. выделены первичные зоны зрительного, слухового, кинестетического восприятия. В норме амплитуда альфа-ритма отражает состояние готовности к повышению функциональной активности данной области коры мозга.
Следовательно, принимаем во внимание распределение максимумов спектральной мощности альфа-ритма у испытуемого в состоянии спокойного бодрствования при закрытых глазах и определяем частоту альфа-ритма, характерную для каждого центрального анализатора органов чувств.
На рис. 2 видно, что усреднённые в группе испытуемых максимумы спектральной мощности альфа-диапазона распределились по зонам сенсорного представительства в соответствии с их функциональным значением. Альфа-ритм с максимальной мощностью над сенсомоторной корой имеет более низкую частоту 7–9 Гц, чем альфаритм с максимальной мощностью в затылочной области коры над первичной зоной зрительного восприятия 9–13 Гц.
В височных зонах первичного слухового восприятия у отдельных индивидуумов в состоянии покоя увеличивалась мощность не только на частоте 12–13 Гц, но и на частоте 7–8 Гц. Возможно, это связано с наличием двух потоков акустических сигналов и раздельных центров восприятия. Первый – сформированный благодаря воздушной проводимости звука через наружный
слуховой проход, барабанную перепонку и цепь слуховых косточек; второй – благодаря тканевой проводимости через ткани черепа.
Для подтверждения принадлежности альфа-ритма определённой частоты к передаче сигналов соответствующего вида ощущений была определена локализация эквивалентных дипольных источников (ЭДИ) происхождения альфа-колебаний в афферентных системах нервных путей и в промежуточных ядрах, через которые проходят сигналы выбранной сенсорной модальности.
На рис. 3 представлены результаты анализа на примере полученных данных испытуемого И. Проведено сопоставление локализации ЭДИ альфа-ритма в глубинных структурах мозга с распределением спектральной мощности альфа-ритма на поверхности коры. Рисунки 3.1 и 3.2 показывают, что альфа-ритм частотой 7–8–9 Гц имеет максимальную спектральную мощность в центральной области коры мозга, где представлены первичные зоны соматосенсорного восприятия – поля 1, 2, 3 по Бродману [15]. ЭДИ этого альфа-ритма локализуются там же, преимущественно в первичных полях по Бродману 1, 2, 3, но они также распределяются по ходу афферентных сигналов в ядрах таламуса и в лимбической области коры мозга в полях по Бродману 23, 25, 27 и 30, 31, которые тесно связаны с развитием эмоций и исполнением инстинктивного поведения. Более высокая плотность ЭДИ наблюдается в ядрах таламуса – основных структурах переключения потоков сенсомоторных сигналов. Нужно подчеркнуть, что речь идёт об ЭДИ происхождения альфа-волн одинаковой частоты 7–8–9 Гц как в центре соматосенсорного восприятия в коре мозга, так и в глубинных структурах, через которые в кору поступают афферентные сигналы той же соматосенсорной модальности. Это может означать, что вся система каналов передачи соматосенсорных сигналов работает в диапазоне 7–8–9 Гц.
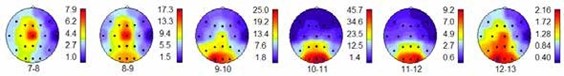
Рис. 2. Результаты группового анализа спектральной мощности альфа-ритма. Максимум мощности ритма 7–9 Гц наблюдается в сенсомоторных зонах, ритма 9–12 Гц – в зрительных зонах коры. Первичные зоны слухового восприятия в височных областях имеют повышенную мощность альфа-ритма на частоте 12–13 Гц. Справа от карты коры мозга – шкала величин мкВ2 спектральной мощности альфа-ритма, внизу – частота альфа-ритма в Гц. (Карту коры мозга и распределение электродов для регистрации ЭЭГ см. на рис. 1)
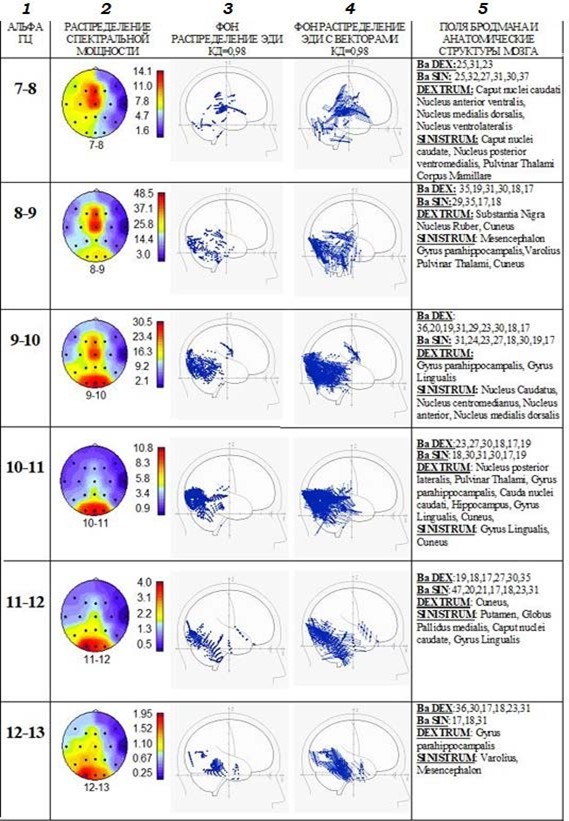
Рис. 3. Испытуемый И. Обозначения столбцов: 1 – частота анализируемого альфа-ритма; 2 – пример распределения спектральной мощности альфа-ритма на поверхности мозга; 3 – локализации ЭДИ происхождения альфа-волн; 4 – локализация тех же ЭДИ с векторами; 5 – анатомическое обозначение локализации ЭДИ
Важно обратить внимание на направленность векторов ЭДИ, локализованных в глубинных структурах мозга: эти вектора направлены в сторону центральных областей коры с сенсомоторными зонами первичного восприятия и этим обуславливают повышение спектральной мощности именно в этих зонах [14].
Рассмотрение спектральных характеристик альфа-ритма частотой 9–10–11–12 Гц позволяет получить убедительные сведения в пользу преимущественного распределения максимумов в затылочной области коры. ЭДИ этого ритма, особенно на частоте 9–10–11 Гц, локализуются в повышенном количестве в первичных и вторичных зонах зрительного восприятия (в полях по Бродману 17, 18, 19), включая Gyrus Lingualis и Cuneus. Они также широко представлены в лимбической системе, в структурах основания мозга, которые отвечают за эмоциональный контроль и запоминание. А также ЭДИ этого ритма распределяются в структурах системы восходящей неспецифической активации ствола мозга и Mesencephalon. В данной системе зрительного восприятия каналы передачи зрительных сигналов работают в том же частотном диапазоне альфа-ритма. В глубинных структурах вектора ЭДИ альфа-ритма частотой 9–10–11–12 Гц в основном направлены к поверхности зрительной коры.
Несколько иначе (рис. 3.2) представлена активность альфа-ритма частотой 12–13 Гц. Распределение спектральной мощности этого ритма показывает максимум не только в зрительной коре, но и в височной области, где расположены зоны первичного слухового восприятия. Концентрация ЭДИ этого ритма приближена к внутренним структурам височной области. Глубинные ЭДИ расположены в стволовых ядрах системы восходящей неспецифической активации коры и в лимбической системе, контролирующей эмоции и свойства памяти. Вектора ЭДИ направлены к поверхности как височных, так и затылочных зон коры мозга.
С другой стороны, выше было отмечено, что в корковых центрах слухового восприятия наблюдается также увеличение мощности альфа-ритма частотой 7–8 Гц. То, что это происходит в височной области (поле 37 по Бродману), где располагается акустико-гностический сенсорный центр речи (Вернике) слева, показано на рис. 3.2.
Этот центр контролирует речевые процессы и ответственен за понимание речи. ЭДИ этого альфа-ритма (рис. 3.3 и 3.4) локализуются в области задних отделов верхней височной извилины слева. Вектора ЭДИ направлены к поверхности височной коры и способствуют увеличению спектральной мощности альфа-ритма на частоте 7–8 Гц.
Альфа-волны, ЭДИ которых локализуются непосредственно в слуховых центрах в височной области коры и в глубинных структурах, показывают для слухового восприятия два частотных диапазона: 7–8–9 Гц и 12–13 Гц. Вектора ЭДИ в каждом частотном диапазоне направлены к поверхности височной коры.
Итак, можно выделить следующие признаки системной деятельности мозга в процессе сенсорного восприятия той или иной модальности.
- Выраженные соотношения между частотой альфа-ритма и модальностью сигналов. Альфа-ритм частотой 7–9 Гц относится к соматосенсорному, 9–12 Гц – к зрительному, 12–13 Гц и 7–9 Гц – к слуховому восприятию.
- ЭДИ альфа-ритма, локализованные в первичной сенсорной зоне коры, по ходу афферентных путей, в структурах лимбической области и восходящей неспецифической активации, объединяются в систему каналов передачи сенсорных сигналов отдельной модальности в едином частотном диапазоне.
- ЭДИ альфа-ритма, объединённые в систему каналов, имеют направления векторов преимущественно в сторону первичного центра сенсорного восприятия той же модальности.
- При высоком уровне дипольности размерность электрических полей ЭДИ достаточна, чтобы достичь поверхности коры. Кроме того, отмечено [14], что соотношения глубинных и поверхностных скальповых потенциалов Uглуб/Uсклп для альфа-ритма наиболее оптимальны и составляют только 1,2-1,5/1.Напротив,длядельта-итета-активности они составляют от 4/1 до 18/1 и даже до 80/1, что затрудняет обнаружение дельта- и тета-волн глубинного происхождения на поверхности коры. По всей вероятности, эфаптическое распространение и тонические влияния альфа-колебаний в среде мозга не только обеспечивают повышение возбудимости ассоциативных путей для передачи информации из одной области в другую, но и способствуют формированию специфической активности с воссозданием внутренних образов.
- Частотная специфичность каналов передачи сенсорных сигналов для каждой модальности может быть обусловлена стробированием – выделением интервала на шкале
частот для увеличения вероятности обнаружения полезных сигналов на фоне помех [8, 9]. Функция стробирования необходима в процессе сканирования с помощью электрических альфа-волн [6, 7], которые могут распространяться непосредственно в средах мозга, влияя на состояние возбудимости мембран отдельных элементов нейронных сетей.
- Эквивалентные дипольные источники происхождения альфа-ритма ЭЭГ с высоким коэффициентом дипольности (КД=>0,98) рассматриваются как наиболее активные локальные зоны, запускающие последовательное перемещение волновых процессов альфа-колебаний. Эти колебания разности электрических потенциалов оказывают непосредственное влияние на организацию первичного восприятия в центральных анализаторах и могут сопровождать сканирование и стробирование с переносом информации в ассоциативные зоны построения сложных образов.
Заключение
Такимобразом,былинетолькоподтверждены известные данные о зонах первичного восприятия коры мозга, но и определены конкретные функциональные параметры рассматриваемых нейросистем в виде частотного стробирования в диапазоне альфаритма, которые могут быть использованы как алгоритмический инструмент для изучения более сложных нейрофизиологических процессов. Предполагается, что этот инструмент может быть применим для получения объективных данных о свойствах сознания человека, эмоциональном состоянии и о некоторых психофизиологических функциях.
Данное исследование проводилось в рамках проекта «EMUSE.Neuro» (г. Москва, Лаборатория Хаоса «XaosLab» – изучение и прогнозирование поведенческих закономерностей хаотических потоков и нейроалгоритмов эмоционального интеллекта, руководитель и инициатор проекта – Александр Кузяев).
МЕЖДУНАРОДНЫЙ ЖУРНАЛ ПРИКЛАДНЫХ И ФУНДАМЕНТАЛЬНЫХ ИССЛЕДОВАНИЙ №7, 2022